Channelrhodopsin - Channelrhodopsin - Wikipedia
Kanalhodopsinlar ning bir oilasi retiniliden oqsillari (rodopsinlar ) vazifasini bajaradi nurli ionli kanallar.[1] Ular xizmat qiladi sezgir fotoreseptorlar bitta hujayrali yashil suv o'tlari, nazorat qilish fototaksis: nurga javoban harakat.[2] Boshqa organizmlarning hujayralarida ifodalangan, ular yorug'likni boshqarishga imkon beradi elektr qo'zg'aluvchanligi, hujayra ichidagi kislota, kaltsiy oqim va boshqa uyali jarayonlar (qarang optogenetika ). Model organizmdan Channelrhodopsin-1 (ChR1) va Channelrhodopsin-2 (ChR2) Chlamydomonas reinhardtii birinchi topilgan kanalradopsinlardir. Variantlar boshqa alg turlaridan klonlangan va yana ko'p narsalar kutilmoqda.
Tuzilishi
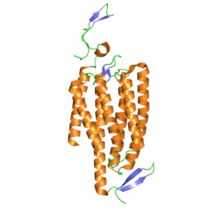
Kanal rodopsinlari tuzilishi jihatidan retiniliden oqsillari. Ular etti transmembran oqsillari kabi rodopsin va yorug'lik izomerizatsiyalanadigan tarkibiga kiradi xromofor hamma-trans -setchatka (an aldegid lotin ning A vitamini ). Retinal xromofor oqsilning qolgan qismi bilan kovalent ravishda protonlangan holda bog'lanadi Shiff bazasi. Aksariyat 7 transmembranli oqsillar G oqsillari bilan bog'langan retseptorlari orqali boshqa ion kanallarini bilvosita ochadigan ikkinchi xabarchilar (ya'ni ular metabotropik ) kanalrhodopsinlari to'g'ridan-to'g'ri ion kanallarini hosil qiladi (ya'ni, ular ionotropik ).[4] Bu hujayrali depolarizatsiyani juda tez, mustahkam va bioinjiniring va nevrologiya dasturlari, shu jumladan foydali qiladi fotostimulyatsiya.
Funktsiya

Tabiiy ("yovvoyi turdagi") ChR2 ko'k nurni yutish bilan yutadi va harakat spektri maksimal 480 nm.[5] Qachonkitrans-reinal kompleks a singdiradi foton, bu barchadan konformatsion o'zgarishni keltirib chiqaraditrans 13 yoshgachacis-reinal. Ushbu o'zgarish transmembran oqsiliga yana birini kiritadi va teshikni kamida 6 to ga ochadi. Milisekundlar davomida to'r pardasi yana trans holatiga tushib, teshikni yopadi va ionlar oqimini to'xtatadi.[4] Natopsidinlarning ko'pi o'ziga xos emas kation kanallarni olib boradigan H+, Na+, K+va Ca2+ ionlari. Yaqinda, anion o'tkazuvchi kanalradopsinlar topilgan.[6]
Kanal-rodopsinlar dizaynerlari
Channelrhodopsinlar asosiy vositadir optogenetika. The C-terminali Channelrhodopsin-2 ning oxiri hujayra ichidagi bo'shliqqa tarqaladi va uni almashtirish mumkin lyuminestsent oqsillar kanal funktsiyasiga ta'sir qilmasdan. Bunday termoyadroviy qurilish ChR2 ekspression hujayralarining morfologiyasini tasavvur qilish uchun foydali bo'lishi mumkin.[7][8] Nuqta mutatsiyalar retinani bog'laydigan cho'ntagiga yaqin kanalrhodopsinning biofizik xususiyatlariga ta'sir ko'rsatishi natijasida turli xil vositalar paydo bo'ldi.
Kinetika
Optik faollashuvdan so'ng kanalning yopilishi C128 yoki D156 oqsillari qoldiqlarini mutatsiyalash orqali kechiktirilishi mumkin. Ushbu modifikatsiya natijasida o'ta sezgir kanal-rodopsinlar paydo bo'ladi, ular ko'k nur zarbasi bilan ochilishi va yashil yoki sariq nur zarbasi bilan yopilishi mumkin (Step-funktsiyasi opsinlari).[9][10][11] E123 qoldig'ini mutatsiyalash kanal kinetikasini (ChETA) tezlashtiradi va natijada hosil bo'lgan ChR2 mutantlari neyronlarni 200 Hz gacha ko'tarish uchun ishlatilgan.[12] Umuman olganda, sekin kinetikasi bo'lgan kanalradopsinlar populyatsiya darajasida yorug'likka ko'proq sezgir, chunki ochiq kanallar vaqt o'tishi bilan kam yorug'lik darajalarida ham to'planib boradilar.
Fototok amplituda
H134R va T159C mutantlari ortib borgan toklarni namoyish etadi va T159 va E123 (ET / TC) kombinatsiyasi bir oz kattaroq toklarga va yovvoyi turdagi ChR2 ga qaraganda bir oz tezroq kinetikaga ega.[13] ChR variantlari orasida ChR1 va ChR2 ning ximerasi va nuqta mutanti bo'lgan ChIEF eng katta fototoklarni va eng kam desensitizatsiyani namoyish etadi va yovvoyi turdagi ChR2 ga o'xshash kinetikaga ega.[14]
To'lqin uzunligi
Kimyoviy kanal-rodopsinlar ChR1 va VChR1 dan transmembranli spirallarni birlashtirib ishlab chiqilgan bo'lib, qizil spektral siljishlar bilan (masalan, C1V1 va ReaChR) ChRlarning rivojlanishiga olib keladi.[11][15] ReaChR membranalar savdosini va sutemizuvchilar hujayralarida kuchli ekspressionni yaxshilagan va miya sopi minimal invaziv, transkranial faollashuvida ishlatilgan. motoneyronlar. Boshqa organizmlarda gomologik ketma-ketlikni izlash natijasida spektral jihatdan yaxshilangan va kuchliroq qizil siljigan kanalodrpsinlar (Chrimson) paydo bo'ldi.[16] ChR2 bilan birgalikda bu sariq / qizil nurga sezgir kanal-rodopsinlar neyronlarning ikkita populyatsiyasini har xil rangdagi yorug'lik impulslari bilan mustaqil ravishda boshqarishga imkon beradi.[17]
Yosunlardan ko'k siljigan kanal-rodopsin topildi Sherffelia dubia. Membrana savdosi va tezligini yaxshilash uchun bir muncha muhandislikdan so'ng, natijada paydo bo'lgan vosita (CheRiff) 460 nm qo'zg'alishda katta fotosuratlar hosil qildi.[18] U genetik jihatdan kodlangan jRCaMP1b kaltsiy ko'rsatkichi bilan birlashtirilgan [19] OptoCaMP deb nomlangan barcha optik tizimda.[20]
Ion selektivligi
L132C mutatsiyasi (CatCh) kaltsiy uchun o'tkazuvchanlikni oshiradi va juda katta oqimlarni hosil qiladi.[21] E90ni musbat zaryadlangan aminokislota argininiga mutatsiyalashgan kanalrhodopsinni o'ziga xos bo'lmagan kation kanalidan xlor o'tkazuvchi kanalga (ChloC) aylantiradi.[22] Kanal teshigidagi manfiy zaryadlangan qoldiqlarni almashtirish orqali Cl- ning selektivligi yanada yaxshilandi teskari potentsial ko'proq salbiy.[23][24] Tanlangan xlorid o'tkazuvchi kanal Rodopsinlar (iChloC, iC ++, GtACR) ko'k nur bilan yoritilganida hujayra madaniyati va buzilmagan hayvonlarda neyronlarning boshoqlanishini inhibe qiladi.
Ilovalar
Channelrhodopsinlar kabi qo'zg'aluvchan hujayralarda osongina ifodalanishi mumkin neyronlar transfektsiya qilishning turli usullaridan foydalangan holda (virusli) transfektsiya, elektroporatsiya, gen qurol ) yoki transgen hayvonlar. Yorug'likni yutuvchi pigment setchatka aksariyat hujayralarda mavjud (ning umurtqali hayvonlar ) kabi A vitamini, buni amalga oshirish fotostimulyatsiya hech qanday kimyoviy birikma qo'shmasdan neyronlar. Kanalhodopsinlar kashf qilinishidan oldin nevrologlar cheklangan edi yozib olish miyadagi neyronlarning faoliyati va o'zaro bog'liq bu faoliyat o'zini tutishi bilan. Bu qayd etilgan asabiy faoliyatni isbotlash uchun etarli emas sabab bo'lgan bu xatti-harakatlar. Genetik modifikatsiyalangan hujayralar tarmoqlarini yorug'lik bilan boshqarish, deb nomlanuvchi yangi paydo bo'layotgan maydon Optogenetika., endi tadqiqotchilarga sabab ma'lum bir neyron guruhidagi faoliyat o'rtasidagi bog'liqlik va aqliy hodisalar, masalan. Qaror qabul qilish. Xatti-harakatlarning optik nazorati nematodalar, mevali chivinlar, zebrafish va sichqonlarda namoyish etilgan.[25][26] Yaqinda, xlorid o'tkazuvchi kanal Rodopsinlar muhandislik qilingan va tabiatda ham topilgan.[6][22] Ushbu vositalar yordamida hujayralar madaniyati va tirik hayvonlardagi neyronlarning ovozini o'chirish uchun foydalanish mumkin manevrli inhibisyon.[23][24]
Yorug'likning bir nechta ranglaridan foydalanish imkoniyatlarini kengaytiradi optogenetik tajribalar. Moviy nurga sezgir bo'lgan ChR2 va sariq rangli faollashtirilgan xlorli nasos halorhodopsin birgalikda ko'p rangli optik faollashtirish va asab faolligini susaytirishga imkon beradi.[27][28] VChR1 mustamlaka algidan Volvox carteri maksimal darajada 535 nmda yutadi va sariq nurli (580 nm) hujayralarni rag'batlantirish uchun ishlatilgan, ammo VChR1 tomonidan ishlab chiqarilgan fotokarorlar odatda juda kichikdir.[29] Biroq, VChR1-ChR2 duragaylari yordamida ishlab chiqilgan yo'naltirilgan evolyutsiya 560 nm maksimal qo'zg'alishni va 600 nm dan yuqori to'lqin uzunliklarida 50% eng yuqori emirishni namoyish etadi.[15][30]
Yorug'lik bilan stimulyatsiya qilingan lyuminestsent yorliqli ChR2 dan foydalangan holda aksonlar va sinapslar aniqlanishi mumkin.[8] Bu induksiya paytida molekulyar hodisalarni o'rganish uchun foydalidir sinaptik plastika.[31] Transfektsiya qilingan madaniylashtirilgan neyron tarmoqlari robototexnika va boshqaruvda qo'llaniladigan ba'zi bir xatti-harakatlarni bajarish uchun rag'batlantirilishi mumkin.[32] Shuningdek, ChR2 miyaning bir tomonidan ikkinchi tomoniga uzoq masofali bog'lanishlarni xaritalashda va individual neyronlarning dendritik daraxtidagi yozuvlarning fazoviy joylashuvini xaritada ko'rsatishda ishlatilgan.[33][34]
Ichki retinal hujayralardagi ChR2 ni ekspluatatsiya qilish orqali ko'r sichqonlarda ko'rish funktsiyasi qisman tiklanishi mumkin.[35][36] Kelajakda ChR2 tibbiy dasturlarni topishi mumkin, masalan. retinaning degeneratsiyasi shakllarida yoki uchun miyani chuqur stimulyatsiya qilish. Optik koklear implantatlar hayvonlar tajribalarida yaxshi ishlashi ko'rsatilgan va birinchi qo'llanilishiga olib kelishi mumkin optogenetika odamlarda.[37][38][39]
Tarix
Mikroalglarning harakatchanligi va fotorentatsiyasi (fototaksis ) yuz yildan oshiq vaqt mobaynida dunyoning ko'plab laboratoriyalarida o'rganilgan.
1980 yilda Ken Foster yosun ko'zlari faoliyati to'g'risida birinchi izchil nazariyani ishlab chiqdi.[40] Shuningdek, u nashr etilgan harakat spektrlarini tahlil qildi va ko'r hujayralarni retinal va retinal analoglari bilan to'ldirdi, bu esa xlorofitlarda harakatlanish reaktsiyalari uchun fotoreseptor degan xulosaga keldi. rodopsin.[41]
Xlorofitlarning fotosuratlari Heamatococcus pluvialis va Chlamydomonas reinhardtii ko'p yillar davomida Oleg Sineshchekov va guruhlarida o'rganilgan Piter Hegemann, natijada 1978 va 1991 yillarda ikkita yarim nashr nashr etildi.[42][43] Harakat spektroskopiyasi va bir vaqtning o'zida fotosuratlar va flagellar urishini yozib olish asosida, fotoreseptor oqimlari va keyingi flagellar harakatlari rodopsin va nazorat fototaksis va fotofobik reaktsiyalar vositasida ekanligi aniqlandi. Qisqa yorug'lik porlashidan keyin fotoreseptor oqimining juda tez ko'tarilishi, rodopsin va kanalning oqsil kompleksida yoki hatto bitta oqsil ichida chambarchas bog'langan degan xulosaga keldi.[44][45]
Biroq, rodopsin-fotoreseptorlarni biokimyoviy tozalash ko'p yillar davomida muvaffaqiyatsiz tugadi.
Endilikda kanalrhodopsinlar ChR1 va ChR2 deb nomlangan rodopsinlarning nukleotidlar ketma-ketligi nihoyat keng miqyosda ochildi est ketma-ketlikdagi loyiha C. reinhardtii. Xuddi shu ketma-ketliklarni mustaqil ravishda taqdim etish GenBank uchta tadqiqot guruhlari tomonidan ularning nomlanishi bilan bog'liq chalkashliklar paydo bo'ldi: Ismlar politsiya-3 va politsiya-4 Hegemann guruhi tomonidan dastlabki topshirish uchun ishlatilgan;[46] csoA va csoB Spudich guruhi tomonidan;[2] va akop-1 va akop-2 Takahashi guruhi tomonidan.[47] Ikkala ketma-ketlik a-da bitta komponentli nur bilan faollashtirilgan kation kanallari sifatida ishlashi aniqlandi Ksenopus oositlar va inson buyrak hujayralari (HEK) Georg Nagel, Ernst Bamberg, Peter Hegemann va boshqalar.[1][4]
Ushbu noodatiy xususiyatni ta'kidlash uchun "kanalrhodopsin" nomi paydo bo'ldi va ketma-ketliklar mos ravishda o'zgartirildi. Ayni paytda ularning alg hujayralarida fotoreseptor oqimlarini hosil qilishdagi rollari Oleg Sineshchekov, Kvan-Xvan Jung va Jon Spudich,[2] va Piter Bertold va Piter Hegemann.[48]
2004 yil noyabr oyida, Chjuo-Xua Pan ga hujjat topshirdi Tabiat Channelrhodopsin bilan yuqtirilgan ko'r sichqonlarda ko'rish qobiliyatini tiklash haqida xabar berish,[iqtibos kerak ] ammo qog'oz rad etildi[iqtibos kerak ] va oxir-oqibat nashr etilgan Neyron 2006 yilda.[iqtibos kerak ]
Shu bilan birga, 2005 yilda uchta guruh ketma-ket ChR2 ni vosita sifatida yaratdi genetik jihatdan maqsadli optik masofadan boshqarish pulti (optogenetika ) ning neyronlar, asabiy zanjirlar va xatti-harakatlar.
Boshida, Karl Deyzserot laboratoriyasi (2005 yil avgustda chop etilgan maqolada) ChR2 ni boshqarish uchun ishlatilishini namoyish etdi sutemizuvchi neyronlar in vitro, millisekundlar bo'yicha vaqtinchalik aniqlikka erishish (boshoqni kechiktirish nuqtai nazaridan ham, vaqtinchalik titrash nuqtai nazaridan ham).[7] Bu juda muhim topilma edi, chunki birinchi navbatda barcha opsinlar (mikrob va umurtqali hayvonlar) talab qiladi setchatka yorug'likni sezuvchi ko-omil sifatida va sutemizuvchilarning markaziy asab hujayralarida retinaning etarli darajasi bo'lishi aniq emas edi, ammo ular shunday; ikkinchidan, bu kichik bir kanal o'tkazuvchanligiga qaramay, sutemizuvchilar neyronlarini harakat potentsiali chegarasidan yuqori darajada haydash uchun etarli quvvatni ko'rsatdi; va uchinchidan, bu kanalodopodinni birinchi optogenetik vosita ekanligini ko'rsatdi, uning yordamida neyronlarning faoliyati vaqtinchalik aniqligi bilan boshqarilishi mumkin (millisekundlarda). Fotostimulyatsiya uchun avvalgi vosita cHARGe madaniylashtirilgan neyronlarda printsipiallikni isbotladi[49] ammo boshqa guruhlar tomonidan hech qachon ishlatilmadi, chunki u soniyalar tartibida aniq ishlaydi, juda o'zgaruvchan va shaxsni boshqarishga imkon bermaydi harakat potentsiali.
Keyinchalik PG Hegemann va Stefan Herlitze guruhlari tomonidan ChR2 ning faoliyatini boshqarish qobiliyatini tasdiqlovchi ikkinchi tadqiqot nashr etildi. umurtqali hayvonlar neyronlar, bu vaqtda jo'ja orqa miyasida.[50] Ushbu tadqiqot birinchi bo'lib ChR2 optik susturucu, umurtqali hayvonlar bilan bir qatorda ifodalangan rodopsin - bu holda birinchi marta qo'zg'aluvchan hujayralarni faollashtirish va shu ikki vosita yordamida bir vaqtning o'zida jim bo'lishini namoyish qilib, to'qimalarni har xil to'lqin uzunliklarida yoritadi.
Aleksandr Gotschalk va Ernst Bamberg guruhlari (eksperimental etakchi Jorj Nagel ishtirokida), agar ChR2 aniq neyronlarda yoki mushak hujayralarida ifodalangan bo'lsa, bashorat qilinadigan xatti-harakatlarni keltirib chiqarishi, ya'ni buzilmagan hayvonlar asab tizimini boshqarishi mumkin, bu holda umurtqasizlar C. elegans.[51] Bu birinchi bo'lib ChR2 yordamida optogenetik eksperimentda hayvonning xatti-harakatini boshqarish, genetik jihatdan aniqlangan hujayra turini optik masofadan boshqarish moslamasini boshqarish. Garchi ikkala jihat o'sha yilning boshida boshqa bir guruh tomonidan tasvirlangan bo'lsa-da, Miesenbok bilvosita nurli ionli P2X2 kanalini tarqatadigan laboratoriya,[52] bundan buyon to'g'ridan-to'g'ri optik faollashuvning kuchi, tezligi, maqsadga muvofiqligi, foydalanish qulayligi va vaqtinchalik aniqligi tufayli qo'zg'aluvchan hujayralarni genetik maqsadli masofadan boshqarish sohasida hukmronlik qilgan kanalrhodopsin singari mikrobial opsinlar, masalan, qafas kabi tashqi kimyoviy birikmalar talab qilinmadi. ligandlar.[53]
Uning asosiy salbiy tomonlarini engish uchun - kichik bir kanalli o'tkazuvchanlik (ayniqsa barqaror holatda), bitta optimal qo'zg'alish to'lqinining chegarasi (~ 470 nm, ko'k) va tiklanishning nisbatan uzoq davom etishi, yuqorida neyronlarning nazorat ostida otilishiga yo'l qo'ymaslik 20-40 Hz - ChR2 yordamida optimallashtirilgan gen muhandisligi. A nuqta mutatsiyasi H134R (mahalliy oqsilning 134 holatidagi histidin aminokislotasini argininga almashtirishi) barqaror holatni o'tkazuvchanligini oshirdi, 2005 yilgi maqolada tasvirlanganidek, ChR2 ni optogenetik vosita sifatida yaratdi. C. elegans.[51] 2009 yilda, Rojer Tsien Laboratoriya ChR2-ni barqaror holatdagi o'tkazuvchanlikni yanada oshirish va ChR1 va ChR2 ximeralarini yaratish va o'ziga xos aminokislotalarni mutatsiyalashtirib, ChEF va CHIEF hosil qilish orqali desensitizatsiyani keskin kamaytirish uchun optimallashtirdi, bu esa 100 Gts gacha bo'lgan harakat potentsiali poezdlarini haydashga imkon berdi.[14][54] 2010 yilda Hegemann va Deisseroth guruhlari mahalliy ChR2 ga E123T mutatsiyasini kiritdilar va ChETA hosil qildilar, u tezroq va o'chiriladi.kinetika, 200 Gts gacha bo'lgan chastotalarda (tegishli hujayralar turlarida) individual harakat potentsialini boshqarishga ruxsat berish.[12][14]
Hegemann va Deisseroth guruhlari, shuningdek, C128S nuqta mutatsiyasining kiritilishi natijasida hosil bo'lgan ChR2-hosilasini qadam funktsiyali vositaga aylantirishini aniqladilar: ko'k chiroq bilan "yoqilganda", ChR2 (C128S) u yoqilguncha ochiq holatda qoladi. sariq nur bilan o'chirilgan - vaqtinchalik aniqlikni yomonlashtiradigan modifikatsiya, lekin yorug'lik sezgirligini ikki darajaga oshiradi.[9] Shuningdek, ular VChR1 ni ko'p hujayrali suv o'tlarida kashf etdilar va tavsifladilar Volvox carteri. VChR1 faqat kichik fotoelektrlarni hosil qiladi, ammo assimilyatsiya spektri ChR2 ga nisbatan qizil tomonga siljiydi.[29] ChR1 ketma-ketligining qismlaridan foydalanib, keyinchalik ikkita neyronal populyatsiyani ikkita to'lqin uzunligida qo'zg'atishga imkon berish uchun fototok amplituda yaxshilandi.[11]
Deisseroth guruhi genetik maqsadli masofadan boshqarish kabi jonli hayvonlarda ko'plab dasturlarni yaratgan kemiruvchilar jonli ravishda,[55] kemiruvchilarda ta'limning optogenetik induktsiyasi,[56] ning eksperimental davolash Parkinson kasalligi kalamushlarda,[57][58] va bilan birikmasi FMRI (opto-fMRI).[59] Boshqa laboratoriyalar barcha optik tajribalar uchun ChR2 stimulyatsiyasini kaltsiy ko'rish bilan birlashtirishga kashshof bo'lishdi,[8] uzoq masofani xaritalash[33] va mahalliy[60] asab zanjirlari, transgen lokusdan ChR2 ifodasi - to'g'ridan-to'g'ri[61] yoki ichida Cre-lox shartli paradigma[60] - shuningdek ikki foton individual hujayralarni faollashtirishga imkon beradigan ChR2 ning qo'zg'alishi.[62][63][64]
2013 yil mart oyida miya mukofoti (Grete Lundbeck miya tadqiqotlari bo'yicha Evropa mukofoti) birgalikda "optogenetikani ixtiro qilgani va takomillashtirgani" uchun Bamberg, Boyden, Deyzserot, Hegemann, Miesenbok va Nagelga berildi.[65] Xuddi shu yili, Hegemann va Nagel ularni oldi Tibbiyot bo'yicha Lui-Jantet mukofoti "kanalodopsinni kashf qilish" uchun. 2015 yilda Boyden va Deisseroth ushbu sovg'ani oldilar Hayot fanlari bo'yicha mukofot va 2020 yilda Miesenbok, Hegemann va Nagel ushbu mukofotlarni oldi Hayot fanlari va tibbiyot sohasidagi shou mukofoti optogenetikani rivojlantirish uchun.
Adabiyotlar
- ^ a b Nagel G, Ollig D, Fuhrmann M, Kateriya S, Musti AM, Bamberg E, Hegemann P (iyun 2002). "Channelrhodopsin-1: yashil suv o'tlarida nurli protonli kanal". Ilm-fan. 296 (5577): 2395–8. doi:10.1126 / science.1072068. PMID 12089443.
- ^ a b v Sineshchekov OA, Jung KH, Spudich JL (iyun 2002). "Chlamydomonas reinhardtii-da ikkita rodopsin fototaksisni past va yuqori zichlikdagi nurga vositachilik qiladi". Proc. Natl. Akad. Ilmiy ish. AQSH. 99 (13): 8689–94. doi:10.1073 / pnas.122243399. PMC 124360. PMID 12060707.
- ^ Kato HE, Zhang F, Yizhar O, Ramakrishnan C, Nishizawa T, Xirata K, Ito J, Aita Y, Tsukazaki T, Hayashi S, Hegemann P, Maturana AD, Ishitani R, Deisseroth K, Nureki O (fevral 2012). "Rodopsin nurli kationli kanal kanalining kristalli tuzilishi". Tabiat. 482 (7385): 369–74. doi:10.1038 / nature10870. PMC 4160518. PMID 22266941.
- ^ a b v Nagel G, Szellas T, Xun V, Kateriya S, Adeishvili N, Bertold P, Ollig D, Hegemann P, Bamberg E (Noyabr 2003). "Channelrhodopsin-2, to'g'ridan-to'g'ri nurli kation-selektiv membranali kanal". Proc. Natl. Akad. Ilmiy ish. AQSH. 100 (24): 13940–5. doi:10.1073 / pnas.1936192100. PMC 283525. PMID 14615590.
- ^ Bamann C, Kirsch T, Nagel G, Bamberg E (yanvar 2008). "Channelrhodopsin-2 fototsiklining spektral xarakteristikalari va uning kanal funktsiyasiga ta'siri". J. Mol. Biol. 375 (3): 686–94. doi:10.1016 / j.jmb.2007.10.072. PMID 18037436.
- ^ a b Govorunova E.G., Sineshchekov OA, Janz R, Lyu X, Spudich JL (2015). "Tabiiy nurli anion kanallari: rivojlangan optogenetika uchun mikrobial rodopsinlar oilasi". Ilm-fan. 349 (6248): 647–650. doi:10.1126 / science.aaa7484. PMC 4764398. PMID 26113638.
- ^ a b Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K (sentyabr 2005). "Milisaniyalik vaqt o'lchovi, asabiy faoliyatni genetik maqsadli optik boshqarish". Nat. Neurosci. 8 (9): 1263–8. doi:10.1038 / nn1525. PMID 16116447.
- ^ a b v Chjan YP, Oertner TG (2007 yil fevral). "Nurga sezgir kanal yordamida sinaptik plastisitni optik induksiyasi". Nat. Usullari. 4 (2): 139–41. doi:10.1038 / nmeth988. PMID 17195846.
- ^ a b Berndt A, Yizhar O, Gunaydin LA, Hegemann P, Deisseroth K (fevral 2009). "Ikki tomonlama barqaror asab tugmachalari". Nat. Neurosci. 12 (2): 229–34. doi:10.1038 / nn.2247. PMID 19079251.
- ^ Schoenenberger P, Gerosa D, Oertner TG (2009). "Zudlik bilan erta genlarni induksiyasini vaqtincha boshqarish". PLOS ONE. 4 (12): e8185. doi:10.1371 / journal.pone.0008185. PMC 2780714. PMID 19997631.
- ^ a b v Yizhar O, Fenno LE, Prigge M, Shnayder F, Devidson TJ, O'Shea DJ, Sohal VS, Goshen I, Finkelshteyn J, Paz JT, Stehfest K, Fudim R, Ramakrishnan C, Gugenard JR, Hegemann P, Dayserot K ( 2011 yil sentyabr). "Axborotni qayta ishlash va ijtimoiy disfunktsiyadagi neokortikal qo'zg'alish / inhibisyon balansi". Tabiat. 477 (7363): 171–8. doi:10.1038 / tabiat10360. PMC 4155501. PMID 21796121.
- ^ a b Gunaydin LA, Yizhar O, Berndt A, Sohal VS, Deisseroth K, Hegemann P (mart 2010). "Ultrafast optogenetik nazorat". Nat. Neurosci. 13 (3): 387–92. doi:10.1038 / nn.2495. PMID 20081849.
- ^ Berndt A, Schoenenberger P, Mattis J, Tye KM, Deisseroth K, Hegemann P, Oertner TG (may 2011). "Kam yorug'lik darajasida tez neyronlarni stimulyatsiya qilish uchun yuqori samaradorlikli kanaloplastiklar". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 108 (18): 7595–600. doi:10.1073 / pnas.1017210108. PMC 3088623. PMID 21504945.
- ^ a b v Lin JY (2011 yil yanvar). "Kanal-rodopsin variantlari bo'yicha foydalanuvchi qo'llanmasi: xususiyatlari, cheklovlari va kelajakdagi o'zgarishlar". Eksperimental fiziologiya. 96 (1): 19–25. doi:10.1113 / expphysiol.2009.051961. PMC 2995811. PMID 20621963.
- ^ a b Lin JY, Knutsen PM, Myuller A, Kleinfeld D, Tsien RY (oktyabr 2013). "ReaChR: kanalrhodopsinning qizil siljigan varianti chuqur transkranial optogenetik qo'zg'alishni ta'minlaydi". Tabiat nevrologiyasi. 16 (10): 1499–508. doi:10.1038 / nn.3502. PMC 3793847. PMID 23995068.
- ^ Klapoetke NC, Murata Y, Kim SS, Pulver SR, Birdsey-Benson A, Cho YK, Morimoto TK, Chuong AS, Carpenter EJ, Tian Z, Vang J, Xie Y, Yan Z, Zhang Y, Chow BY, Surek B, Melkonian M, Jayaraman V, Konstantin-Paton M, Vong GK, Boyden ES (mart 2014). "Alohida nerv populyatsiyalarining mustaqil optik qo'zg'alishi". Tabiat usullari. 11 (3): 338–46. doi:10.1038 / nmeth.2836. PMC 3943671. PMID 24509633.
- ^ Hooks BM, Lin JY, Guo C, Svoboda K (mart 2015). "Ikki kanalli sxemani xaritalash asosiy motor korteksida sensorimotor konvergentsiyani aniqlaydi". Neuroscience jurnali. 35 (10): 4418–26. doi:10.1523 / JNEUROSCI.3741-14.2015. PMC 4355205. PMID 25762684.
- ^ Hochbaum DR, Zhao Y, Farhi SL, Klapoetke N, Verli, CA, Kapoor V, Zou P, Kralj JM, Maclaurin D, Smedemark-Margulies N, Saulnier JL, Boulting GL, Straub C, Cho YK, Melkonian M, Vong GK, Harrison DJ, Murthy VN, Sabatini BL, Boyden ES, Campbell RE, Cohen AE (avgust 2014). "Sutemizuvchi neyronlarda muhandislik mikrobial rodopsinlari yordamida to'liq optik elektrofiziologiya". Tabiat usullari. 11 (8): 825–33. doi:10.1038 / nmeth.3000. PMC 4117813. PMID 24952910.
- ^ Dana H, Mohar B, Sun Y, Narayan S, Gordus A, Xasseman JP, Tsegaye G, Xolt GT, Xu A, Valpita D, Patel R, Maklin JJ, Bargmann CI, Ahrens MB, Shrayter ER, Jayaraman V, Looger LL , Svoboda K, Kim DS (2016 yil mart). "Nerv faolligini ko'rish uchun sezgir qizil oqsil kaltsiy ko'rsatkichlari". eLife. 5. doi:10.7554 / eLife.12727. PMC 4846379. PMID 27011354.
- ^ Afshar Saber V, Gasparoli FM, Dirks MG, Gunn-Mur FJ, Antkowiak M (2018). "Biologik asabiy tarmoqlarni o'rganish bo'yicha barcha optik tahlillar". Nevrologiya chegaralari. 12: 451. doi:10.3389 / fnins.2018.00451. PMC 6041400. PMID 30026684.
- ^ Kleinlogel S, Feldbauer K, Dempski RE, Fotis H, Wood PG, Bamann C, Bamberg E (aprel 2011). "Ca² + o'tkazuvchan kanalli Rodopsin CatCh bilan ultra nurga sezgir va tez neyron faollashuvi" (PDF). Tabiat nevrologiyasi. 14 (4): 513–8. doi:10.1038 / nn.2776. PMID 21399632.
- ^ a b Vietek J, Viegert JS, Adeishvili N, Shnayder F, Vatanabe H, Tsunoda SP, Vogt A, Elstner M, Oertner TG, Hegemann P (aprel 2014). "Kanal Rodopsinni nurli xlorli kanalga aylantirish". Ilm-fan. 344 (6182): 409–12. doi:10.1126 / science.1249375. PMID 24674867.
- ^ a b Wietek J, Beltramo R, Scanziani M, Hegemann P, Oertner TG, Wiegert JS (oktyabr 2015). "In vivo jonli neyronlarning faolligini inhibe qilish uchun yaxshilangan xlor o'tkazuvchan kanalrhodopsin". Ilmiy ma'ruzalar. 5: 14807. doi:10.1038 / srep14807. PMC 4595828. PMID 26443033.
- ^ a b Berndt A, Li SY, Vietek J, Ramakrishnan C, Shtaynberg EE, Rashid AJ, Kim H, Park S, Santoro A, Frankland PW, Iyer SM, Pak S, Erlund-Rixter S, Delp SL, Malenka RC, Xosselin SA, Carlén M, Hegemann P, Deisseroth K (yanvar 2016). "Optogenetikaning strukturaviy asoslari: kanaloplastik ionlarining selektivligini aniqlash omillari". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 113 (4): 822–9. doi:10.1073 / pnas.1523341113. PMC 4743797. PMID 26699459.
- ^ Duglass AD, Kraves S, Deisseroth K, Schier AF, Engert F (avgust 2008). "Zebrafish somatosensor neyronlarida bitta kanalli rodopsin-2 tomonidan uyg'otilgan boshoqlar natijasida paydo bo'lgan qochish harakati". Curr. Biol. 18 (15): 1133–7. doi:10.1016 / j.cub.2008.06.077. PMC 2891506. PMID 18682213.
- ^ Xuber D, Petreanu L, Gitani N, Ranade S, Hromadka T, Meynen Z, Svoboda K (yanvar 2008). "Bochka korteksidagi siyrak optik mikrostimulyatsiya erkin harakatlanuvchi sichqonlardagi harakatlarni o'rganadi". Tabiat. 451 (7174): 61–4. doi:10.1038 / nature06445. PMC 3425380. PMID 18094685.
- ^ Xan X, Boyden ES (2007). "Ko'p rangli optik faollashtirish, sustlashish va asab faolligini desinxronizatsiya qilish, bir pog'onali vaqtinchalik rezolyutsiya bilan". PLOS ONE. 2 (3): e299. doi:10.1371 / journal.pone.0000299. PMC 1808431. PMID 17375185.
- ^ Zhang F, Vang LP, Brauner M, Liewald JF, Kay K, Watzke N, Wood PG, Bamberg E, Nagel G, Gottschalk A, Deisseroth K (aprel 2007). "Nerv sxemalarining multimodal tezkor optik so'rovi". Tabiat. 446 (7136): 633–9. doi:10.1038 / nature05744. PMID 17410168.
- ^ a b Zhang F, Prigge M, Beyrière F, Tsunoda SP, Mattis J, Yizhar O, Hegemann P, Deisseroth K (iyun 2008). "Qizil siljigan optogenetik qo'zg'alish: Volvox carteri dan kelib chiqqan holda tezkor neyronlarni boshqarish vositasi". Nat. Neurosci. 11 (6): 631–3. doi:10.1038 / nn.22020. PMC 2692303. PMID 18432196.
- ^ Yizhar O, Fenno LE, Prigge M, Shnayder F, Devidson TJ, O'Shea DJ, Sohal VS, Goshen I, Finkelshteyn J, Paz JT, Stehfest K, Fudim R, Ramakrishnan C, Gugenard JR, Hegemann P, Dayserot K ( 2011 yil iyul). "Axborotni qayta ishlash va ijtimoiy disfunktsiyadagi neokortikal qo'zg'alish / inhibisyon balansi". Tabiat. 477 (7363): 171–8. doi:10.1038 / nature10360. PMC 4155501. PMID 21796121.
- ^ Chjan YP, Xolbro N, Oertner TG (2008 yil avgust). "Yagona sinapslarda plastisitni optik induktsiya qilish alfaCaMKII ning o'ziga xos birikmasini aniqlaydi". Proc. Natl. Akad. Ilmiy ish. AQSH. 105 (33): 12039–44. doi:10.1073 / pnas.0802940105. PMC 2575337. PMID 18697934.
- ^ Xu Z, Ziye X, Kreyg H, Silviya F (2013 yil dekabr). Spike-ga asoslangan bilvosita trening, asabiy tarmoq orqali boshqariladigan virtual hasharot. IEEE Qarori va nazorati. 6798-6805-betlar. CiteSeerX 10.1.1.671.6351. doi:10.1109 / CDC.2013.6760966. ISBN 978-1-4673-5717-3.
- ^ a b Petreanu L, Xuber D, Sobchik A, Svoboda K (may 2007). "Channelrhodopsin-2 yordami bilan uzoq masofali kallozal proektsiyalarning sxemasini xaritalash". Nat. Neurosci. 10 (5): 663–8. doi:10.1038 / nn1891. PMID 17435752.
- ^ Petreanu L, Mao T, Sternson SM, Svoboda K (2009 yil fevral). "Neokortikal qo'zg'atuvchi aloqalarning subcellular tashkiloti". Tabiat. 457 (7233): 1142–5. doi:10.1038 / nature07709. PMC 2745650. PMID 19151697.
- ^ Bi A, Cui J, Ma YP, Olshevskaya E, Pu M, Dizhoor AM, Pan ZH (2006 yil aprel). "Mikrobial tipdagi rodopsinni tashqi ifodasi fotoreseptor degeneratsiyasi bo'lgan sichqonlarda vizual reaktsiyalarni tiklaydi". Neyron. 50 (1): 23–33. doi:10.1016 / j.neuron.2006.02.026. PMC 1459045. PMID 16600853.
- ^ Lagali PS, Balya D, Avatramani GB, Myunx TA, Kim DS, Busskamp V, Cepko CL, Roska B (iyun 2008). "ON bipolyar hujayralariga yo'naltirilgan nurli faol kanallar retinaning degeneratsiyasida ko'rish funktsiyasini tiklaydi". Nat. Neurosci. 11 (6): 667–75. doi:10.1038 / nn.2117. PMID 18432197.
- ^ Hernandez VH, Gehrt A, Reuter K, Jing Z, Jeschke M, Mendoza Schulz A, Hoch G, Bartels M, Vogt G, Garnham CW, Yawo H, Fukazawa Y, Augustine GJ, Bamberg E, Kygler S, Salditt T, de Hoz L, Strenzke N, Moser T (2014 yil fevral). "Eshitish yo'lining optogenetik stimulyatsiyasi". J Clin Invest. 124 (3): 1114–29. doi:10.1172 / JCI69050. PMC 3934189. PMID 24509078.
- ^ Mager T, Lopez de la Morena D, Senn V, Schlotte J, D'Errico A, Feldbauer K, Wrobel C, Jung S, Bodensiek K, Rankovic V, Browne L, Huet A, Jüttner J, Wood PG, Letzkus JJ, Moser T, Bamberg E (may, 2018). "Yuqori chastotali qizil pog'onali optogenetika orqali yuqori chastotali asab pog'onasi va eshitish signalizatsiyasi". Nat Commun. 9 (1): 1750. doi:10.1038 / s41467-018-04146-3. PMC 5931537. PMID 29717130.
- ^ Keppeler D, Martins Merino R, Lopez de la Morena D, Bali B, Huet AT, Gehrt A, Wrobel C, Subramanian S, Dombrowski T, Wolf F, Rankovic V, Neef A, Moser T (2018). "Optimallashtirilgan Chronosni nishonga olish yo'li bilan eshitish yo'lining ultrafast optogenetik stimulyatsiyasi". EMBO J. 37 (24): e99649. doi:10.15252 / embj.201899649. PMC 6293277. PMID 30396994.
- ^ Foster KW, Smith R (1980). "Fototaktik suv o'tlarida engil antennalar". Mikrobiologik sharhlar. 44 (4): 572–630. PMC 373196. PMID 7010112.
- ^ Foster KW, Saranak J, Patel N, Zarilli G, Okabe M, Kline T, Nakanishi K (oktyabr 1984). "Rodopsin - bu bir hujayrali eukaryot Chlamydomonasdagi fototaksis uchun funktsional fotoreseptor". Tabiat. 311 (5988): 489–491. doi:10.1038 / 311756a0. PMID 6493336.
- ^ Litvin FF, Sineshchekov OA, Sineshchekov VA (1978). "Haematococcus pluvialis alga fototaksisidagi fotoreseptor elektr potentsiali". Tabiat. 271 (5644): 476–478. doi:10.1038 / 271476a0. PMID 628427.
- ^ Xarz H, Hegemann P (iyun 1991). "Rhodopsin bilan boshqariladigan kaltsiy oqimlari Xlamidomonalarda". Tabiat. 351 (6326): 489–491. doi:10.1038 / 351489a0.
- ^ Holland EM, Braun FJ, Nonnengässer C, Harz H, Hegemann P (Fevral 1996). "Xlamidomonalardagi rodopsin bilan qo'zg'atilgan fotokarorlarning tabiati. I. kinetika va ikki valentli ionlarning ta'siri". Biofiz. J. 70 (2): 924–931. doi:10.1016 / S0006-3495 (96) 79635-2. PMC 1224992. PMID 8789109.
- ^ Braun FJ, Hegemann P (1999 yil mart). "Yashil suv o'tlari Volvox carteri ko'zida ikkita nurli faol o'tkazuvchanlik". Biofiz. J. 76 (3): 1668–1778. doi:10.1016 / S0006-3495 (99) 77326-1. PMC 1300143. PMID 10049347.
- ^ Kateriya, S. Fuhrmann, M. Hegemann, P .: To'g'ridan-to'g'ri yuborish: Chlamydomonas reinhardtii retinal biriktiruvchi oqsil (cop4) geni; GenBank kirish raqami AF461397
- ^ Suzuki T, Yamasaki K, Fujita S, Oda K, Iseki M, Yoshida K, Watanabe M, Daiyasu H, Toh H, Asamizu E, Tabata S, Miura K, Fukuzawa H, Nakamura S, Takahashi T (Fevral 2003). "Xlamidomonalardagi arxeologik tipdagi rodopsinlar: model tuzilishi va hujayra ichidagi lokalizatsiya". Biokimyo. Biofiz. Res. Kommunal. 301 (3): 711–7. doi:10.1016 / S0006-291X (02) 03079-6. PMID 12565839.
- ^ Berthold P, Tsunoda SP, Ernst OP, Mages V, Gradmann D, Hegemann P (iyun 2008). "Channelrhodopsin-1 zudlik bilan nurli depolarizatsiya orqali xlamidomonalarda fototaksis va fotofobik reaktsiyalarni boshlaydi". O'simlik hujayrasi. 20 (6): 1665–1677. doi:10.1105 / tpc.108.057919. PMC 2483371. PMID 18552201.
- ^ Zemelman BV, Li GA, Ng M, Miesenbok G (yanvar 2002). "Genetik jihatdan o'zgargan neyronlarning selektiv fotostimulyatsiyasi". Neyron. 33 (1): 15–22. doi:10.1016 / S0896-6273 (01) 00574-8. PMID 11779476.
- ^ Li X, Gutierrez DV, Xanson MG, Xan J, Mark MD, Chiel H, Hegemann P, Landmesser LT, Herlitze S (dekabr 2005). "Omurgalı rodopsin va yashil suv o'tlari kanalrhodopsin tomonidan asabiy va tarmoq faolligini tez invaziv bo'lmagan faollashishi va inhibatsiyasi". Proc. Natl. Akad. Ilmiy ish. AQSH. 102 (49): 17816–21. doi:10.1073 / pnas.0509030102. PMC 1292990. PMID 16306259.
- ^ a b Nagel G, Brauner M, Livald JF, Adeishvili N, Bamberg E, Gottschalk A (dekabr 2005). "Kanrhodopsin-2 ning Caenorhabditis elegans qo'zg'atuvchi hujayralarida nurlanish bilan faollashishi tezkor xatti-harakatlarni keltirib chiqaradi". Curr. Biol. 15 (24): 2279–84. doi:10.1016 / j.cub.2005.11.032. PMID 16360690.
- ^ Lima SQ, Miesenbok G (aprel 2005). "Neyronlarning genetik maqsadli fotostimulyatsiyasi orqali xatti-harakatlarni masofadan boshqarish". Hujayra. 121 (1): 141–52. doi:10.1016 / j.cell.2005.02.004. PMID 15820685.
- ^ Jang F, Vang LP, Boyden ES, Deisseroth K (oktyabr 2006). "Channelrhodopsin-2 va qo'zg'aluvchan hujayralarni optik boshqarish". Nat. Usullari. 3 (10): 785–92. doi:10.1038 / nmeth936. PMID 16990810.
- ^ Lin JY, Lin MZ, Shtaynbax P, Tsien RY (mart 2009). "Rivojlangan kanallar va kinetikasi bilan takomillashtirilgan kanaloplastik variantlarining tavsifi". Biofiz. J. 96 (5): 1803–14. doi:10.1016 / j.bpj.2008.11.034. PMC 2717302. PMID 19254539.
- ^ Adamantidis AR, Zhang F, Aravanis AM, Deisseroth K, de Lecea L (noyabr 2007). "Gipokretin neyronlarining optogenetik boshqaruvi bilan zondlangan uyg'onishning asabiy substratlari". Tabiat. 450 (7168): 420–4. doi:10.1038 / nature06310. PMC 6744371. PMID 17943086.
- ^ Tsai HC, Zhang F, Adamantidis A, Stuber GD, Bonci A, de Lecea L, Deisseroth K (may, 2009). "Dopaminerjik neyronlarning fazali otilishi o'zini tutishi uchun etarli". Ilm-fan. 324 (5930): 1080–4. doi:10.1126 / science.1168878. PMC 5262197. PMID 19389999.
- ^ Gradinaru V, Mogri M, Tompson KR, Xenderson JM, Deisseroth K (aprel 2009). "Parkinsoniy asab tizimining optik dekonstruksiyasi". Ilm-fan. 324 (5925): 354–9. CiteSeerX 10.1.1.368.668. doi:10.1126 / science.1167093. PMC 6744370. PMID 19299587.
- ^ Kravitz AV, Freeze BS, Parker PR, Kay K, Thwin MT, Deisseroth K, Kreitzer AC (iyul 2010). "Bazal ganglionlar sxemasini optogenetik boshqarish orqali parkinsoniyalik harakatlanishlarni tartibga solish". Tabiat. 466 (7306): 622–6. doi:10.1038 / nature09159. PMC 3552484. PMID 20613723.
- ^ Lee JH, Durand R, Gradinaru V, Zhang F, Goshen I, Kim DS, Fenno LE, Ramakrishnan C, Deisseroth K (iyun 2010). "Neytronlar tomonidan boshqariladigan global va mahalliy FMRI signallari turi va simlari bo'yicha optogenetik jihatdan aniqlanadi". Tabiat. 465 (7299): 788–92. doi:10.1038 / nature09108. PMC 3177305. PMID 20473285.
- ^ a b Kätzel D, Zemelman BV, Buetfering C, Völfel M, Miesenbok G (yanvar 2011). "Neokortikal qo'zg'atuvchi hujayralar bilan inhibitor birikmalarning ustunli va laminali tashkil etilishi". Nat. Neurosci. 14 (1): 100–7. doi:10.1038 / nn.2687. PMC 3011044. PMID 21076426.
- ^ Vang H, Peca J, Matsuzaki M, Matsuzaki K, Noguchi J, Qiu L, Vang D, Zhang F, Boyden E, Deisseroth K, Kasai H, Hall WC, Feng G, Augustine GJ (may 2007). "Channelrhodopsin-2 transgen sichqonlarida fotostimulyatsiya yordamida sinaptik ulanishning yuqori tezlikda xaritasi". Proc. Natl. Akad. Ilmiy ish. AQSH. 104 (19): 8143–8. doi:10.1073 / pnas.0700384104. PMC 1876585. PMID 17483470.
- ^ Mohanty SK, Reinscheid RK, Liu X, Okamura N, Krasieva TB, Berns MW (oktyabr 2008). "Infraqizil lazer mikro nurlari bilan ikki fotonli qo'zg'alish yordamida yuqori fazoviy rezolyutsiyaga ega kanalrhodopsin 2-sezgir qo'zg'atuvchi hujayralarni chuqur faollashtirish". Biofiz. J. 95 (8): 3916–26. doi:10.1529 / biofhysj.108.130187. PMC 2553121. PMID 18621808.
- ^ Rickgauer JP, Tank DW (sentyabr 2009). "Kanal Rodopsin-2 ning ikki fotonli qo'zg'alishi to'yinganlikda". Proc. Natl. Akad. Ilmiy ish. AQSH. 106 (35): 15025–30. doi:10.1073 / pnas.0907084106. PMC 2736443. PMID 19706471.
- ^ Andrasfalvy BK, Zemelman BV, Tang J, Vaziri A (iyun 2010). "Ikki fotonli bitta hujayrali haykalning nurlari bilan neyronlarning faolligini optogenetik boshqarish". Proc. Natl. Akad. Ilmiy ish. AQSH. 107 (26): 11981–6. doi:10.1073 / pnas.1006620107. PMC 2900666. PMID 20543137.
- ^ Reiner A, Isacoff EY (oktyabr 2013). "Brain Prize 2013: optogenetika inqilobi". Nörobilimlerin tendentsiyalari. 36 (10): 557–60. doi:10.1016 / j.tins.2013.08.005. PMID 24054067.
Qo'shimcha o'qish
- Hegemann P. (2008). "Algal sensorli fotoreseptorlari". Annu Rev Plant Biol. 59: 167–189. doi:10.1146 / annurev.arplant.59.032607.092847. PMID 18444900. (Kanal rodopsinlari va boshqa fotoreseptorlarning Naturel funktsiyasi yashil rangda)
- Arenkiel BR, Peca J, Davison IG va boshq. (2007 yil aprel). "Kanal rodopsin-2 ni ifodalovchi transgen sichqonlardagi in-vivo jonli nurlanish ta'sirida faollashuv". Neyron. 54 (2): 205–18. doi:10.1016 / j.neuron.2007.03.005. PMC 3634585. PMID 17442243. (Transgen sichqonlarda kanalodopsin yordamida miya konturini o'rganish)
- Bi A, Cui J, Ma YP va boshq. (2006 yil aprel). "Mikrobial tipdagi Rodopsinning ektopik ifodasi sichqonlardagi fotoreseptorlar degeneratsiyasi bilan vizual javoblarni tiklaydi". Neyron. 50 (1): 23–33. doi:10.1016 / j.neuron.2006.02.026. PMC 1459045. PMID 16600853. (Ko'rni davolash uchun potentsial kanalrhodopsinni qo'llash)
Tashqi havolalar
- OpenOptogenetics.org, optogenetika haqida keng qamrovli wiki.
- Optogenetik resurs markazi / Deisseroth laboratoriyasi
- Boyden laboratoriyasi
- Zhuo-Xua Pan laboratoriyasi
- Hegemann laboratoriyasi
- Brain mukofoti 2013 yil optogenetika ixtirosi uchun